
Bryozoans in the English Chalk

Dr Paul D Taylor (UK)
We are very fortunate in Britain to host one of the most remarkable deposits in the entire geological record, the Chalk. The Late Cretaceous Chalk (with a capital ‘C’) is an extremely pure limestone, famous for the White Cliffs of Dover and responsible for the landscape of rolling hills and dry valleys, forming the ‘downs’ and ‘wolds’ that stretch through England from Devon in the southwest, to Yorkshire in the northeast. The economic importance of the Chalk to the early human inhabitants of Britain was enormous because the flints contained within it could be fashioned into axe heads and hard cutting tools.
Why is the Chalk so special geologically? It is a rare example of a pelagic sediment – an open ocean sediment – that was deposited over the continental shelf. This occurred at a time when global sea-level was high and the supply of terrigenous clastic sediment into the sea was minimal. The Chalk is an oceanic ooze composed mainly of the disaggregated plates – coccoliths – of coccolithophores, planktonic microalgae with exquisitely engineered skeletons of calcite. Unfathomable numbers of coccolithophores sank to the seabed over a period of some 35 million years to produce the thick accumulation of Chalk that today extends over northern Europe and into western Asia.
The Chalk is a favourite hunting ground for fossil collectors, yielding beautifully preserved specimens, especially of echinoids. But closer inspection of the Chalk shows that the dominant macrofossils are often bryozoans. These colony-forming invertebrates are not only present in great abundance, but are also very diverse (Taylor, 2002a), probably exceeding a thousand species in the English Chalk alone.
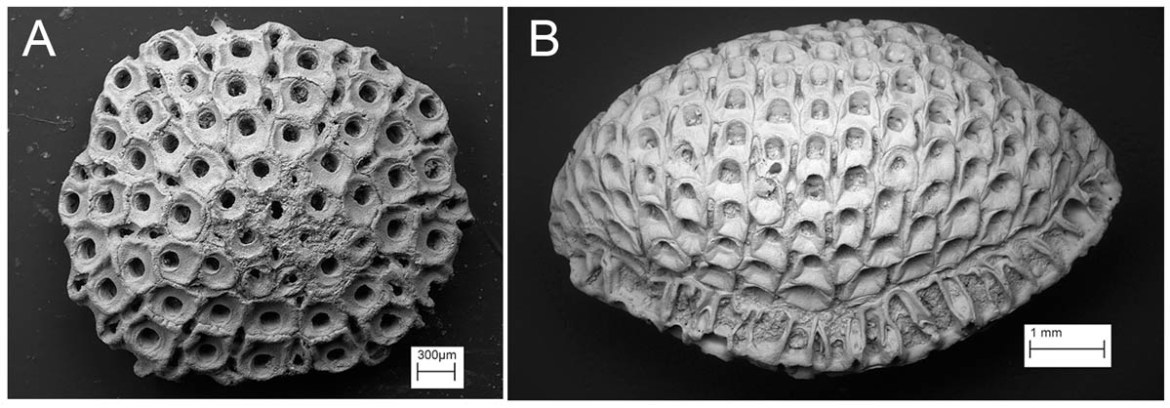
When I started work almost 40 years ago as a bryozoan researcher at the Natural History Museum, London (then called the British Museum (Natural History)), I was struck by the fact that almost one-third of the drawers in the fossil bryozoan collection were occupied by specimens from the Chalk. Many of the Chalk bryozoans in this huge collection encrust the irregular echinoids Echinocorys (Fig. 1) and Micraster, some undoubtedly acquired incidentally from collectors, who decided to pass their ‘imperfect’ echinoids to the bryozoan curator rather than attempting to scrape-off the offending bryozoans. In fact, a very high proportion of Echinocorys tests from the British Chalk are encrusted by bryozoans – just check your own collection.

In addition to the encrusting bryozoans, the Chalk is a rich source of broken branches from bush- or palm-like bryozoan colonies. A lot of these bryozoans in the NHMUK were collected during the nineteenth and early twentieth centuries at a time when the countryside was dotted with small chalk pits available to avid enthusiasts. Prominent among the band of Chalk aficionados were Arthur Rowe (1858–1926), a medical doctor who devised the early macrofossil zonation of the Chalk, and Christopher Gaster (1878–1963), an energetic, non-vocational geologist, who published several important papers on the Chalk of southern England.
Working as a warder at Chatham Prison, William Gamble also sold a large number of specimens to the museum (Fig. 1). It is easy, though perhaps entirely fanciful, to imagine Gamble ordering the prisoners under his charge to break-up boulders of Chalk so that he could search for bryozoans and other fossils. Solicitor Reginald Marr Brydone (1873-1943) was another collector of Chalk bryozoans who, unlike the others mentioned here, published numerous papers, describing in all more than 400 new species of bryozoans. Brydone’s massive collection of Chalk bryozoans and other fossils went to the Sedgwick Museum in Cambridge.
How to collect and recognise Chalk bryozoans
Bryozoans are generally scarce in the Lower Chalk (Cenomanian stage), but become commoner in the Upper Chalk (Coniacian to Maastrichtian stages). The cliffs of Sussex, Kent and Norfolk are better hunting grounds than those of Yorkshire, where the Chalk is harder and generally less fossiliferous.
As already mentioned, bryozoans in the Chalk frequently encrust echinoids, so a good way of finding them is to collect irregular echinoids. After cleaning with nothing more severe than a toothbrush, the surfaces of the echinoids should be examined using a hand lens or a binocular microscope. Some of the bryozoans appear as almost circular patches, usually less than a centimetre in diameter (Fig. 2A), whereas others have branching colonies that spread out more widely over the tests (Fig. 2B). Additional substrates worth inspecting for encrusting bryozoans are inoceramid bivalve shells and belemnite guards, although these tend to be less productive than echinoid tests.

Fragments of erect bryozoans can be found on air-weathered surfaces of Chalk where they typically appear as narrow stick-like branches less than 2mm in diameter, or as slightly broader plate-like pieces. When the chalk is uncemented and the bryozoans are sufficiently abundant, a bulk sample of sediment can be taken to be sieved for bryozoans. Outside Britain, the Chalk can be so packed with bryozoans that it is almost a bryozoan limestone. This is true of the youngest Chalk beneath the Cretaceous/Paleocene boundary in Denmark, as well as some of the Chalk exposed in the cliffs near Étretat, on the northern coast of France. In southern England, Arthur Rowe obtained delicate bryozoans from ‘flint meals’, the very soft chalk occasionally found inside flint nodules (Fig. 3).

Silicified bryozoans can also be seen when flints are broken open or may be preserved standing out from the surface of some flint nodules (Fig. 4).

As with bryozoans from other geological deposits, Chalk bryozoans have a modular construction, each of the modules (zooids) forming the colony being a millimetre or less in size and having an opening through which the feeding apparatus (lophophore) was protruded during life. In cheilostome bryozoans, the zooids have box-like skeletons, whereas in cyclostome bryozoans, zooidal skeletons are essentially tubular in shape.
Identifying Chalk bryozoans to species or genus level can be daunting for the non-specialist. This is in part due to the lack of modern publications comprehensively describing and illustrating these fossils, but also because of difficulties caused by the evolution of colonies of deceptively similar superficial appearance in numerous unrelated bryozoan groups. Furthermore, some of the differences between species are extremely subtle and even examination using a good quality optical microscope may fail to show the critical features. These difficulties are compounded by the whiteness of the bryozoans against the white chalk matrix – it was once customary to stain Chalk bryozoans with red or blue watercolour paint or ink to emphasise their key features. Nowadays, a scanning electron microscope is essential for the scientific study of Chalk bryozoans and most of the images in this article were made using such an instrument.
Colony forms
Colonial animals tend to occur in a great variety of shapes and sizes, and bryozoan colonies are no exception. Three basic colony types can be found in the Chalk: encrusting, erect and free-living. As mentioned above, encrusting bryozoans are usually either sheet-like patches, with the constituent zooids tightly spaced leaving no gaps in the colony, or branching, the zooids being arranged in single file (‘runners’) or with a few across the width of each branch (‘ribbons’). A considerable diversity of genera of cheilostomes and cyclostomes in the Chalk develop sheet-like colonies, far too many to be covered fully here. The common, sheet-like cheilostome species formerly placed in the extant genus Membranipora have now been transferred to such genera as Wilbertopora (Fig. 5A), Marginaria (Fig. 5B) and Pyriporella (Fig. 5C), depending on the distribution of spines (or their bases) and defensive zooids called avicularia.

Likewise, cyclostome sheets once placed in Berenicea have been moved to Hyporosopora, Plagioecia (Fig. 6A) and other genera based on the shape of the reproductive zooids (gonozooids), or to Diplosolen for species having smaller zooids (nanozooids) interspersed between the normal-sized feeding zooids (Fig. 6B). Actinopora is another cyclostome sheet (Fig. 6C), but, in this genus, the apertures of the zooids are arranged in radiating rows, superficially resembling the septa of a solitary coral.

The commonest runner-like cheilostome in the Chalk is Herpetopora, a genus characterised by zooids with long, narrow ‘tails’ (Fig. 7), while Stomatopora and Voigtopora (Fig. 2B) are typical cyclostome runners.

Erect bryozoan colonies are usually preserved broken into branch fragments. It is rare to find them still attached to the substrate on which they were anchored, although the bases of erect colonies (Fig. 8) with branches snapped-off can sometimes be found on echinoid tests and so on.

Occasionally, almost intact colonies are found: a couple of examples are shown here, one from the collection of Arthur Rowe has been carefully prepared from the soft chalk matrix (Fig. 9A). The other is the type specimen (Fig. 9B) of Homoeosolen ramulosus, described in 1850 by William Lonsdale, curator of the Geological Society in London. One of the most distinctive of bryozoans found in the Chalk is another species described by Lonsdale, Siphoniotyphlus plumatus (Fig. 9C).

This cyclostome bryozoan, which is often found in flints, has flattened branches, with the apertures of the feeding zooids arranged in arcs diverging from the centre of the branch. At the edges of the branches are narrow, kenozooids lacking apertures. Cheilostome bryozoans belonging to the genus Rhagasostoma occur both as encrusting and erect colonies. The latter have strap-like branches. At high magnification, it is possible to see the distinctive defensive zooids (avicularia) of Rhagasostoma, which in most species have scimitar-like curved ends (Fig. 10).

Free-living bryozoans in the Chalk comprise cheilostomes of two types. Lunulites has small, cap-shaped colonies, with the zooids opening on the gently convex upper surface of the colony (Fig. 11A). In life, Lunulites colonies would have supported themselves above the seabed using the hair-like appendages of special zooids called vibracula. While bryozoans closely similar to Chalk Lunulites can still be found in modern seas, the second type of free-living Chalk bryozoan is without modern analogues.
Resembling a fusuline foraminiferan, Volviflustrellaria taverensis has small, rugby ball- to spindle-shaped colonies, with zooids opening over the entire surface (Fig. 11B). It is thought that colonies rolled along the seabed, the axis of the spindle oriented parallel to the seabed. It is not known whether they were self-propelled – as are some colonies of living bryozoans resembling Lunulites – were rolled along by their own growth that altered the balance of the colony, or by currents or even the activities of mobile animals on the Chalk seafloor.
Cribrimorphs
The most intricate and beautiful bryozoans found in the Chalk are cribrimorph cheilostomes. The Late Cretaceous was the hey-day for cribrimorphs, although this group is still represented in modern seas. The defining feature of cribrimorphs is the so-called costate frontal wall of the feeding zooids. This is formed from spines that are bent horizontally to overarch the frontal surface, meeting along the mid-line of the zooid and often linked by cross-connections along their lengths (Fig. 12).

The spines can contain small pores, and the structure of the zooid is further complicated in some genera by the development of polymorphic zooids that partly overgrow the costate frontal wall and may form elaborate extensions around the aperture. Most Chalk cribrimorphs have encrusting colonies and although less common than other groups of encrusting cheilostomes, they are very diverse.
William Dickson Lang (1878–1966), one of my predecessors at the Natural History Museum, made a detailed study of Cretaceous cribrimorphs, introducing a wealth of new species, genera and families. Known also for his research on the stratigraphy of the Lias of Dorset, Lang was an advocate of the theory of orthogenesis (Taylor, 2002b). Briefly, this proposes that evolutionary lineages follow pre-defined trends, which generally culminate in their extinction as features that were once useful became disadvantageous.
The classic example of orthogenesis leading to extinction was considered to be the Irish Giant Deer (Megaloceros giganteus), with its huge antlers believed to have prevented the animals from holding their heads up. With respect to Cretaceous cribrimorphs, Lang recognised a series of evolutionary lineages, each beginning with simple forms having modestly calcified zooids and leading to complex species with thickly calcified zooids.
According to Lang, in the final species belonging to each lineage, the zooids secreted an excess of calcium carbonate and quite literally entombed themselves in their own coffins, leading to the extinction of the lineage. Lang’s theory of cribrimorph orthogenesis won little support, even at a time when the processes driving evolution were still being debated. One of his critics was Reginald Brydone, who had personal experience of cribrimorph bryozoans. Brydone was particularly antagonistic towards Lang (Taylor et al, 2018), disagreeing with him about the ‘correct’ way to describe and illustrate bryozoans and various other matters.
Conclusions
Despite the great abundance of bryozoans in the English Chalk, we remain remarkably ignorant about the exact number of species present and their stratigraphical ranges, let alone what they can tell us about the changing palaeoenvironments during the long period of deposition of the Chalk. And Lang’s detailed research on cribrimorph bryozoans that led to his ideas of orthogenesis has yet to be fully revised. How did bryozoans adapt to the conditions on the Chalk seabed, and how did they evolve in this peculiar and demanding environment? There is immense scope for new discoveries and interpretations to unlock the secrets of these ubiquitous Chalk fossils.
Further reading
Fossils of the Chalk: Guide, Palaeontological Association No 2 (2nd edition), edited by Andrew B Smith and David J Batten, The Palaeontological Association, London (2002), 374 pages (Paperback), ISBN: 0901702781
Logging the Chalk, by Rory N Mortimore, Whittles Publishing, Caithness, Scotland (2014). 357 pages, (hardback), ISBN: 978-184995-098-5
The Chalk of Sussex and Kent, Geologists’ Association Guide No 57, by Rory N Mortimore, The Geologists’ Association, London (1997), 139 pages (Paperback), ISBN: 0-780900717833
References
Taylor, P.D. 2002a. Bryozoans. Pp. 53–75 in Smith, A.B. & Batten, D.J. (eds). Fossils of the Chalk. Second Edition. Palaeontological Association, London.
Taylor, P.D. 2002b. W.D. Lang, orthogenesis and the evolution of Cretaceous cribrimorph Bryozoa. Pp. 275–298 in Wyse Jackson, P.N. & Spencer Jones, M.E. (eds). Annals of Bryozoology. International Bryozoology Association, Dublin.
Taylor, P.D., Mackinlay, H. & Pethers, H. 2018. Reginald Marr Brydone (1873–1943) and his scientific animosity with William Dickson Lang (1878–1966). Pp. 151–190 in Wyse Jackson, P.N. & Spencer Jones, M. E. (eds). Annals of Bryozoology 6. International Bryozoology Association, Dublin.