
Mammoths in the freezer

Adrian Lister (UK)
As palaeontologists, we are used to relying on the preserved hard parts of extinct organisms – shells, bones, teeth and so on – to reconstruct their appearance and adaptations in life. The reconstruction of soft tissue relies upon our knowledge of related living forms, plus clues such as the scars of muscle attachments on bones or shells. Exceptions include body outlines preserved in the fine-grained sediments of Lagerstätte, such as in the Eocene of Messel (Germany) or the Cambrian Burgess Shale (Canada); or, even more rarely, organisms preserved in 3D, of which the most familiar source is Tertiary amber.
Among mammals, the most celebrated case of exceptional preservation is provided by the carcasses preserved in permafrost in Siberia (Russia), Alaska (USA) and the Yukon (Canada), at localities lying almost exclusively north of the Arctic Circle (Lister and Bahn, 2007). Almost all date to the last glaciation, with radiocarbon dates typically in the range 50 to 10,000 years ago. Species from which partial or whole carcasses have been recovered include bison, horse, wolverine, woolly rhinoceros and, above all, the woolly mammoth. The reason for the preponderance of these is unclear, although it may partly be a matter of reporting bias, other species being considered less interesting or less valuable when discovered by local people.
Even so, not more than a dozen or so complete or largely complete mammoth carcasses have been recovered to date. While Siberian natives have doubtless been finding these remains for millennia, the first carcass to be scientifically studied was excavated in 1806 (Fig. 1) and brought back to St Petersburg, where it still stands in the Zoology Museum.

However, by the time it was recovered, much of the flesh had decayed or been eaten by wolves – the specimen, known as the Adams mammoth after its excavator, is now a skeleton with some patches of skin and muscle remaining on the face and elsewhere (Fig. 2). In 1901, the Beresovka carcass was discovered, with much of its flesh intact, but it was encased in frozen ground and its extraction involved lighting fires around it in an attempt to melt the ice, and finally hacking it out in pieces. On the mounted skin in St Petersburg, one can still see the lines where it was stitched back together.

The majority of recovered finds have been made since the 1970s. Recovered in 1972, the Shandrin mammoth was remarkable for having 291kg of food still in its intestines, one of several individuals for which we have directly preserved evidence of diet (see below). Many of the other recovered individuals have been juveniles of various ages. The male baby mammoth nicknamed ‘Dima’, discovered in 1977, was the most complete individual known up to that time and the first to be subject to detailed scientific study, with internal organs and preservation of structure down to the cellular level.
Finds, or at least reported finds, seem to have accelerated in recent years, with at least five carcasses having come to light since 2007. This is probably due to a combination of increasing awareness of the value of the remains by local people; the finding of carcasses as a by-product of the increasingly active search for mammoth ivory; and the longer annual thaw of permafrost, as a result of global climate warming. Each new discovery still makes headlines, sometimes for spurious reasons.
One of the more recent finds, a juvenile nicknamed ‘Yuka’ (Fig. 3), had a large area of skin and part of the skeleton missing from its back, leading to suggestions that the animal had been killed and filleted by prehistoric hunters. It now seems likely that this was accidental damage by the excavation machine. In a carcass discovered in 2013, initial reports claimed ‘running blood’. More detailed analysis has indeed revealed the remains of blood in its veins, but coagulated, with partly degraded blood cells.

The most complete carcass discovered to date is the baby mammoth Lyuba, which is currently on display at the Natural History Museum in London until 7 September 2014 (Fig. 4). Her external preservation is breathtaking – the skin is textured and folded just like that of an elephant, while delicate structures, such as her trunk tip and female genitalia, are perfectly preserved. She has been the subject of the most intensive scientific study revealing, for example, her precise age of 35 days by counting microscopic growth lines in her teeth (Rountrey et al, 2012). Other revelations about mammoth biology from Lyuba and other carcasses are described below.

How do the carcasses come to be preserved? Burial in the permanently frozen ground of the far north is clearly the key, but the means by which individuals arrived there seem to have been quite varied. The Beresovka mammoth still had a wad of plant food between its teeth (Fig. 5) and bones that had broken more or less at the time of death.
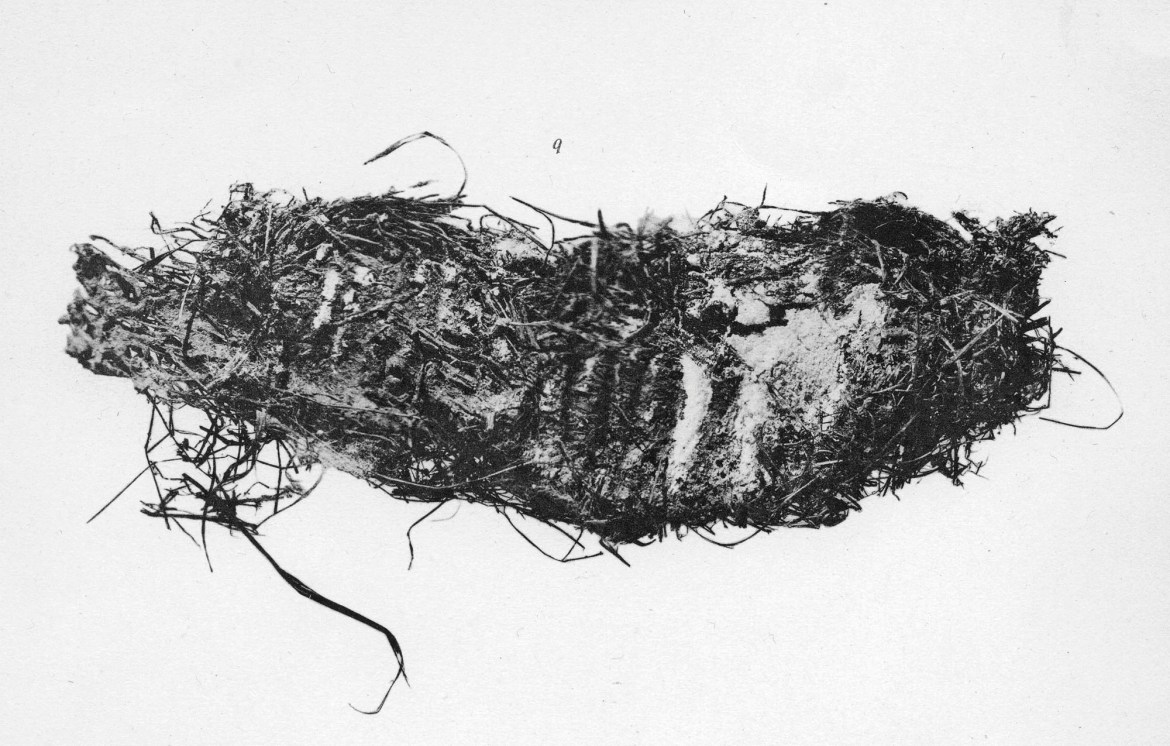
Although, as described above, the geology of the site was scarcely studied, it seems likely that the animal fell into a crevasse (common in the Arctic, due to the seasonal freeze-thaw cycle) and was covered by mobile sediment, which froze around him. Dima is reconstructed as having sunk into soft mud, but with his head exposed, the animal eventually dying of starvation or exhaustion – this scenario is suggested by his emaciated body and tufts of his own hair in his stomach. In the case of Lyuba, death was also due to sinking into soft mud, but was more sudden – CT scans reveal her trunk and upper respiratory tract packed with mud, as she tried to take a last breath.
Dan Fisher, who has led scientific studies on Lyuba, suggests that preservation was initiated by bacteria producing lactic acid, which partly pickled the carcass as it lay in the still-fluid mud (Muller, 2009). Eventually, however, all of the carcasses became encased in icy sediment as the ground around them froze during the winter and, if the surface quickly became covered by more sediment, the carcass found itself at a depth where the spring thaw never reached (Fig. 6).

Over the millennia, the carcasses have become freeze-dried – all their water gradually disappearing by the process of sublimation and forming ice crystals in the surrounding sediment. The end result is akin to the Egyptian mummies – which were deliberately dehydrated by the use of various salts. Therefore, the carcasses do not have to be kept frozen after excavation and are, for the most part, stable at room temperature and humidity.
The frozen carcasses have taught us many things about mammoth biology that we would never have known purely from hard parts (Lister, 2014). For example, several carcasses preserve the ear and/or tail, and these are both much smaller than in living elephants. This presumably was an adaptation to the arctic climate, reducing heat loss and avoiding frostbit in these thin, exposed organs. The coat was thick and covered the entire body – in adults, the guard hairs were up to a metre long, with much finer, shorter underwool next to the skin. A recent study (Tridico et al, 2013) has identified a third layer of intermediate-sized hairs in between.
The same study examined the hairs microscopically and concluded that the commonly-observed orange colour (for example, Fig. 3) was artificial, due to a combination of keratin decay and fungal growth. In some cases, such as in the carcass recovered from the Liakhov Islands in 1901 and now in the Natural History Museum of Paris, the hair is preserved dark brown (Fig. 7). This was probably the original colour of mammoths, although in some areas of the body, the guard hairs seem to have lacked pigment altogether and may have been colourless (Tridico et al, 2014), giving a mottled appearance. The dark colouration has also been corroborated by sequencing DNA from a gene known to affect hair colour in living mammals (Workman et al, 2011).

DNA studies have revealed another adaptation of the woolly mammoth. Its haemoglobin, the protein in red blood cells that transports oxygen, differs slightly from that of living elephants, and tests show that it was better at transferring its oxygen under conditions of low external temperature (Campbell et al, 2010). This remarkable discovery – again, impossible from standard palaeontological observation – was made by actually synthesising mammoth haemoglobin in the laboratory and running it through standard physiological tests as though it were a sample of modern blood.
Further observations on frozen carcasses give us insights into mammoth behaviour. On the Yukagir mammoth, an adult partial carcass discovered in northern Yakutia in Russia, during 2002, the exquisitely-preserved head (Fig. 8) shows a small opening between the eye and ear, demonstrating that mammoths – the males at least – produced secretions from their temporal glands. By analogy with living elephants, these presumably functioned as strong pheromones during an annual ‘musth’ period, when the males would have become highly aggressive and sexually active. The opening is absent in Lyuba, but whether this is because she is a female, or because it had not yet formed in this juvenile, is uncertain.

Several of the individuals preserve food remains in their stomachs, giving us once again an unusually direct insight into the behaviour of an extinct species. The plant remains are usually dominated by grass, but the precise composition varies considerably. One individual had gorged itself on moss in the day or two before its death; another, recently investigated with the new technique of DNA-profiling the range of plants represented, had fed on low herbaceous plants with relatively little grass (Willerslev et al, 2014).
The impression is of animals roaming across the largely treeless glacial environment and feeding on whatever low-growing vegetation they encountered at a particular time. Lyuba’s stomach contained –apart from the remains of her mother’s milk – well-chewed grass that could not have been produce by her hardly-worn baby teeth, leading to the suggestion that, like many herbivorous mammals today, she ate her mother’s dung to acquire the bacteria she would need for digesting plant food (Fisher et al, 2012).
The mammoth’s trunk contained no bones and its structure would have been unknown, but for the frozen remains. It was similar to that of elephants, but with some intriguing differences. The ‘finger and thumb’ projections at the end were longer (Fig. 3) – especially the outer one – and this would have aided the plucking of the predominantly low-growing plants of its diet. The baby mammoth Lyuba (Fig. 4) shows a remarkable structure, only suggested by previously-discovered partial trunks – flanges of skin running the length of the trunk on each side. Its function can only be guessed at – maybe it was used for scooping snow in the winter when there was no liquid water?
More carcasses of mammoth and other ice-age species are bound to be discovered in the future and, together with the further development of techniques (such as DNA analysis and CT scanning), will only increase our understanding of these fascinating creatures.
References
Campbell, K.L., Roberts, J.E.E., Watson, L.N., Stetefeld, J., Sloan, A.M., Signore, A.V., Howatt, J.W., Tame, J.R.H., Rohland, N., Shen, T.-J., Austin, J.J., Hofreiter, M., Ho, C., Weber, R.E. & Cooper, A. 2010. Substitutions in woolly mammoth hemoglobin confer biochemical properties adaptive for cold tolerance. Nature Genetics 42: 536-540.
Fisher, D.C., Tikhonov, A.N., Kosintsev, P. A., Rountrey, A.N., Buigues, B. & van der Plicht, J. 2012. Anatomy, death, and preservation of a woolly mammoth (Mammuthus primigenius) calf, Yamal Peninsula, northwest Siberia. Quaternary International 255: 94-105.
Lister, A. 2014. Mammoths: Ice Age Giants. London: The Natural History Museum.
Lister, A. & Bahn, P. 2007. Mammoths: Giants of the Ice Age. London: Frances Lincoln.
Muller, T. 2009. Ice Baby. National Geographic 215, part 5 (May 2009): 30-49.
Rountrey, A.N., Fisher, D.C., Tikhonov, A.N., Kosintsev, P.A., Lazarev, P.A., Boeskorov, G. & Buigues, B. 2012. Early tooth development, gestation, and season of birth in mammoths. Quaternary International 255: 196-205.
Tridico S.R., Rigby, P., Kirkbride, K.P., Haile, J. & Bunce, M. 2014. Megafaunal split ends: microscopical characterisation of hair structure and function in extinct woolly mammoth and woolly rhino. Quaternary Science Reviews 83: 68-75.
Willerslev, E. et al. 2014. Fifty thousand years of Arctic vegetation and megafaunal diet. Nature 506: 47-51.
Workman, C., Dalen, L., Vartanyan, S., Shapiro, B., Kosintsev, P., Sher, A., Gotherstrom, A., Barnes, I., 2011. Population-level genotyping of coat colour polymorphism in woolly mammoth (Mammuthus primigenius). Quat. Sci. Rev. 30: 2304-2308.